Ecosystem-Level Root Production and Mortality in Response to Climate Change

Dr. Vijayalaxmi Kinhal
April 9, 2026 at 8:12 pm | Updated April 14, 2026 at 2:44 am | 7 min read
- Increasing carbon dioxide produces various root production and root mortality trends in different ecosystems
- Temperature rises increase root production, with diverse effects on root mortality.
- Drought can initially increase root production, but as water deficit intensity and duration increase, it reduces root production and increases root mortality.
- Differences in root quantification methods can make global comparisons and trend analysis challenging.
Knowledge of underground plant root responses to climate change is understandably less than that for above-ground vegetation, due to difficulties in data collection. Therefore, studies on changes in root production, mortality, and turnover at the ecosystem level are few. Climate change is characterized by rising carbon dioxide levels, temperatures, and drought, all of which affect ecosystem-level dynamics. In this article, we summarize the major findings on the effects of these factors on root production and mortality.
Roots Responses are Plastic

Figure 1: Difference in root production in various ecosystems, Ma et al. (2021). (Image credits: https://usys.ethz.ch/en/news-events/news/archive/2021/06/root-to-shoot)
Subscribe to receive our monthly round-up of articles.
Plant roots demonstrate remarkable plasticity in their responses to the environment by altering morphology and function. These changes occur at the individual level but have ecosystem-level impacts.
Though estimates vary, roots collectively make up 24% of plant biomass globally. The portion of biomass underground differs among species and, more broadly, by habit. Thus, roots make up 67% of biomass in grasslands, 47% in shrublands, and 22% in forests; see Figure 1. Since root biomass constitutes a significant portion of plants, it can influence ecosystem-level dynamics. Root influences on plant-level functions have ecosystem-level consequences for the water, nutrient, and carbon cycles, and support soil microbial and fauna. These ecosystem functions can be altered by long-term changes in the roots driven by climate change. Therefore, it is crucial to understand how roots respond to various climate change parameters.
To understand the effects of climate change on ecosystems, the varying functions of different root types should be considered. In forests dominated by trees, woody roots are important as carbon storage organs, while the fine-roots are involved in nutrient and water uptake. Fine-roots can contribute to 60% of carbon cycling in soils. In grasslands, coarse and fine-roots are considered together to study production and turnover. The impact of root turnover also differs between annual and perennial plants. For crops, the role in nutrient uptake is more important than the contribution to carbon flux, which is crucial in natural systems.
Ecosystems such as tropical forests with high diversity will also have a wide range of root responses. It is also important to remember that species-level and ecosystem-level responses will differ.
Climate drivers such as elevated carbon dioxide (CO2), warming, and drought differ in their pattern of change. CO2 increases gradually and uniformly across the globe, while some regions are warming and others are cooling. Similarly, drought due to climate change is more common in Australia, southern Africa, South America, and some parts of North America; see Figure 2.
Roots have been found to respond to major climate change drivers, but the proportion of responses varies by driver and across root categories, as discussed below.
Elevated Carbon Dioxide
The uniform and global rise of CO2 does not have a uniform effect on root production and turnover, according to a review by Norby and Jackson (2000). Photosynthesis was expected to increase, boosting root production. However, several factors, such as ecosystem type, plant lifespan, and nutrient availability, influence the effects of CO2.
Crops
Roots of annual crops are larger and more branched due to elevated CO2, but only when there is sufficient nutrient and water availability; in areas with less nutrient and water availability, the increase in root production is inefficient. There was no change in root turnover in crops, since annual crops experience less mortality over their lifespan. All roots die synchronously at the end of the crop cycle, for example, as observed in wheat.
Grasslands
Changes in root production and turnover are inconsistent, with fewer than 50% of studies reporting increased root growth under elevated CO2. Some areas show no change in production or mortality even after two years of CO2 enrichment; however, root distribution changes. More roots grow in the topsoil layer due to greater soil moisture and CO2.
Forests
The various types of forests, vast areas, and the number of species involved produce a varied trend.
Conifers: In coniferous forests, data collection was possible only in younger seedlings and saplings, and not from the deeper-rooted adults. Among the young conifers, fine-root growth increased due to elevated CO2, but not the root-to-shoot ratio, indicating that larger plants were producing more roots. Moreover, scientists expect root growth to stop over time once it reaches a maximum, just as the canopy reaches a maximum leaf area index (LAI). Although root turnover increases with elevated CO2, the rate is inconsistent. For example, fine-root production increased by 37% and turnover by 26% in a 15-yr Pinus taeda stand in North Carolina, USA, but the relative turnover-to-production ratio did not change significantly. In other cases, however, fine-root mass and mortality in pine trees increased with elevated CO2 only in high-nitrogen soils.
Deciduous: Field experiments show that deciduous trees are more responsive to elevated CO2. Fine-root density, that is, mass per unit area, increases by 60-140% in elevated CO2. The increase in fine-root production exceeded the increase in LAI, indicating a specific effect on roots. Fine-root mortality was also higher. For example, in separate experiments in an Acer saccharum–Acer rubrum stand in the USA and in Liquidambar styraciflua stands, fine-root production and mortality increased.
This indicates a difference in root response between temperate fertile deciduous hardwood and coniferous stands.
Tropical forests: Coarse roots and fine-root production in tropical forests are responsive and increase with higher CO2 levels. However, the root-to-shoot ratio is inconsistent and varies with the soil nutrient status, especially phosphorus. Elevated CO2 alone reduces or has no effect on the root-to-shoot ratio.
Warming
Generalizations are challenging because roots in different ecosystems have evolved in response to distinct environmental conditions, including air and soil temperatures. Soil temperature rises must also be considered, as they generally increase the nitrogen mineralization rate, benefiting plant production. As warming increases, higher nutrient availability and photosynthesis increase plant biomass, including root biomass. If nutrient and water availability are sufficient, root mortality should not increase, resulting in higher root turnover.
Global warming causes very high temperatures in the tropics, in some cases exceeding the photosynthetic optimum, thereby lowering biomass. However, Yaffar et al. (2024), who synthesized 93 studies across 24 tropical countries, found that higher temperatures increase production and root biomass in the tropics when sufficient nutrients and water are available.
Global warming also induces temporal changes, with earlier spring onset of root and vegetative growth in perennials, shifting timing, and increasing carbon flux.
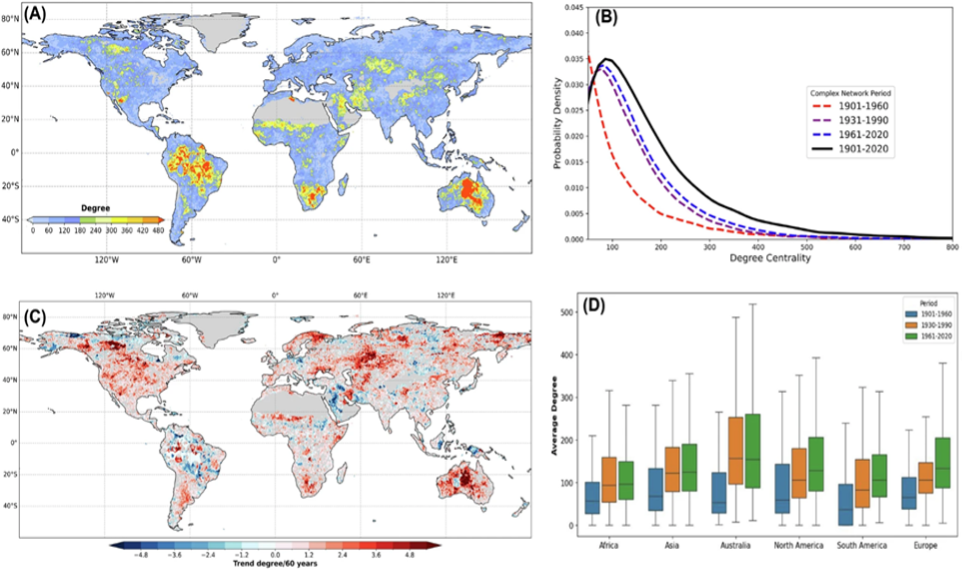
Figure 2: Spatial connectivity structure of global droughts. A Degree centrality of the global drought network, illustrating areas of high spatial clustering. B Degree distribution across different time periods, showing a heavy tail indicative of drought hubs. C Temporal trends in degree centrality (1901-2020), identified via the Mann-Kendall test (p < 0.05). D Box plot of degree centrality for continents across three periods, showing Australia with the highest degree,” Bhatia et al. (2026). (Image credits: https://doi.org/10.1038/s43247-025-03111-5)
Drought
Drought events are expected to become more severe, longer, and more frequent in some areas of the globe, increasing tree mortality. Also, droughts do not occur at the same time or with the same intensity. Only 1.84 to 6.5% of the globe has synchronized areas affected by drought; see Figure 2.
Drought reduces plant growth and survival by reducing photosynthesis and increasing desiccation. Plants close their stomata to reduce water loss, but this also decreases carbon assimilation, reducing biomass.
Initially, when drought starts, plants can increase carbon allocation to roots, thereby increasing root-to-shoot ratios and root length, expanding the soil volume explored for water. However, as drought intensifies or persists, a reduction in photosynthesis stops this reallocation of carbon. The reduction in photosynthesis and increased tissue desiccation, as water acquisition by plants declines, results in less root production. In severe drought conditions, it can lead to plant mortality. Hence, drought reduces root production, while root mortality, especially of fine-roots, increases.
Though fine-root turnover reflects local water availability, at the global scale, drought doesn’t explain fine-root turnover patterns after accounting for temperature impacts. Forests manage to maintain relatively constant fine-root production despite interannual variations in soil moisture, indicating that trees may be more resilient to drought effects than anticipated.
Episodic climate change drivers such as flooding, cyclones, and wildfires can also alter root production and mortality patterns worldwide.
Measuring Root Production and Mortality
In meta-analyses and syntheses that draw on studies from across the globe, scientists acknowledge that, in some cases, differences in root production and mortality may not be due to environmental conditions but to measurement methods. Each method has its advantages and disadvantages. Norby and Jackson (2000) note that minirhizotrons, which can non-destructively quantify growth and mortality, are increasingly used for the purpose. However, they caution that researchers should be aware that root death can be ambiguous and difficult to detect with this method, as mortality occurs gradually and even after decomposition begins, some root functions may continue. CID BioScience Inc. offers two root imagers for use in minirhizotron systems with 105-meter-long root tubes, helping to reach soil depths where fine-roots in trees grow.
Contact us for more information on the CI-600 In-Situ Root Imager and CI-602 Narrow Gauge Root Imager.
Sources
Bhatia, U., Poonia, H., Mansoor Tantary, D. et al. Regional responses to oceanic variability constrain global drought synchrony. Commun Earth Environ 7, 86 (2026). https://doi.org/10.1038/s43247-025-03111-5
Florida State University. (2014, May 5). Global warming not uniform around the globe: Some areas were recently cooling. ScienceDaily. Retrieved March 17, 2026 from www.sciencedaily.com/releases/2014/05/
Hudiburg, T., Mathias, J., Bartowitz, K., Berardi, D. M., Bryant, K., Graham, E., … & Lynch, L. (2023). Terrestrial carbon dynamics in an era of increasing wildfire. Nature Climate Change, 13(12), 1306-1316.
Ma H, Mo L, Crowther TW et al. (2021). The global distribution and environmental drivers of aboveground versus belowground plant biomass. Nat Ecol Evol. Doi: external page 10.1038/s41559-021-01485-1
Norby, R. J., & Jackson, R. B. (2000). Root dynamics and global change: seeking an ecosystem perspective. New Phytologist, 147(1), 3–12. doi:10.1046/j.1469-8137.2000.00676.x
Yaffar, D., Lugli, L. F., Wong, M. Y., Norby, R. J., Addo‐Danso, S. D., Arnaud, M., … & Cusack, D. F. (2024). Tropical root responses to global changes: a synthesis. Global Change Biology, 30(7), e17420.
Related Products
Most Popular Articles
- Transpiration in Plants: Its Importance and Applications
- Leaf Area – How & Why Measuring Leaf Area…
- How to Analyze Photosynthesis in Plants: Methods and Tools
- Plant Respiration: Its Importance and Applications
- The Forest Canopy: Structure, Roles & Measurement
- Stomatal Conductance: Functions, Measurement, and…
- Forest & Plant Canopy Analysis – Tools…
- Root Respiration: Importance and Applications
- 50 Best Universities for Plant Science
- The Importance of Leaf Area Index (LAI) in…



