Roots, Shoots, and Photosynthesis: Measuring Tree Sapling Response to Drought

Scott Trimble
December 5, 2020 at 2:34 am | Updated December 5, 2020 at 2:34 am | 7 min read
Drier and longer summers are taking a toll on trees in temperate regions acclimatized to more rainfall. Therefore, in an attempt to suggest efficient propagation and maintenance practices to match new climate patterns, scientists are studying how various species deal with drought. Advanced analytic tools that can be used consistently in the field are a necessary accessory.
Increasing Dryness and Temperature
Due to climate change, the world is experiencing shorter winters, and longer, hotter summers. The growing trend of the early arrival of spring is causing trees to leaf sooner, which, in turn, leads to higher evapotranspiration. As a result, trees use more soil water and, during summer, with accompanying higher temperatures, deal with drier soils.
As time goes on, warmer regions will experience more drought. The frequency of extreme warm periods expected to increase by a minimum of 73% in Northern Europe but will go up by 173% in Southern and Mediterranean Europe.
Subscribe to receive our monthly round-up of articles.
Therefore, vegetation in the drier Mediterranean is expected to be affected to a greater extent by drought.
The effect on forests is less well-studied; however, trees drying in the wild have raised an alarm, as many species are not only environmentally, but also economically valuable.
Several studies are looking at both the above-ground and below-ground responses of trees to drought. Based on agricultural research, scientists expect trees to invest more biomass to grow deeper and larger roots that will reach more soil water during drought. How resources are allocated depends on nutrient availability and genotype.
Since seedlings and saplings bear the most harmful effects, this article focuses on sapling response to drought and how various precision tools are aiding in their study.
Phenotypic Plasticity Responses in Root Growth of Beech
European beech (Fagus sylvatica L.) is dominant in Central Europe, as it is a strong competitor for resources and grows in a broad range of soil types and rainfall conditions. However, its weakness lies in its relatively high susceptibility to drought.
We know that, due to drought, the beech sheds leaves and experiences canopy dieback, but root responses have, until recently, been left unstudied.
Scientists in Germany sought to understand the effect of drought on fine root production, morphology, and turnover in four beech varieties from different hydrologic conditions.
The researchers also compared the drought response of above and belowground parts, as well as the role of phenotypic plasticity and genetics.
Phenotypic plasticity allows a species to react differently and adapt to variations in environmental conditions; thus, species with high phenotypic plasticity are able to survive in many habitats. Genotypic differences typically result in a species being better adapted to one habitat than others.
The scientists transplanted 25 beech saplings from each habitat to an outdoor Rhizolab. Here, the saplings were grown in long containers. Transparent root tubes, up to 125 cm deep, were distributed to observe root growth and death.
Soil water content (SWC) of 10% was maintained in the well-watered treatment and at 5% SWC in the drought treatment.
Isozyme analysis of ten enzymes was performed for all four varieties. Their genetic diversity, heterozygosity, and genetic distance were also estimated. The genetic distance was considered to be 100% if the genetic structures were completely different and 0% if they were identical.
The mini-rhizotron CI-600 In-Situ Root Imager, manufactured by CID Bio-Science Inc., was used to take root scans across the entire root tube surface once every two weeks, during winter and spring until the summer drought. The CI-600 takes high-resolution images that are analyzed by the accompanying RootSnap software to give root length, area, volume, diameter, and branching angle.
Temporal changes in roots were analyzed by another software, WinRHIZOTron. The disappearance of a root was recorded as its death. The root lifespan was measured as the number of days from root appearance to their disappearance in scans.
In summer, at the end of the experiment, the trees were carefully harvested. Roots were separated according to size, and their dry weight was estimated. For the larger roots, the surface area was also recorded. Leaf size and area were also measured for all leaves in a tree, and dry weight was estimated.]
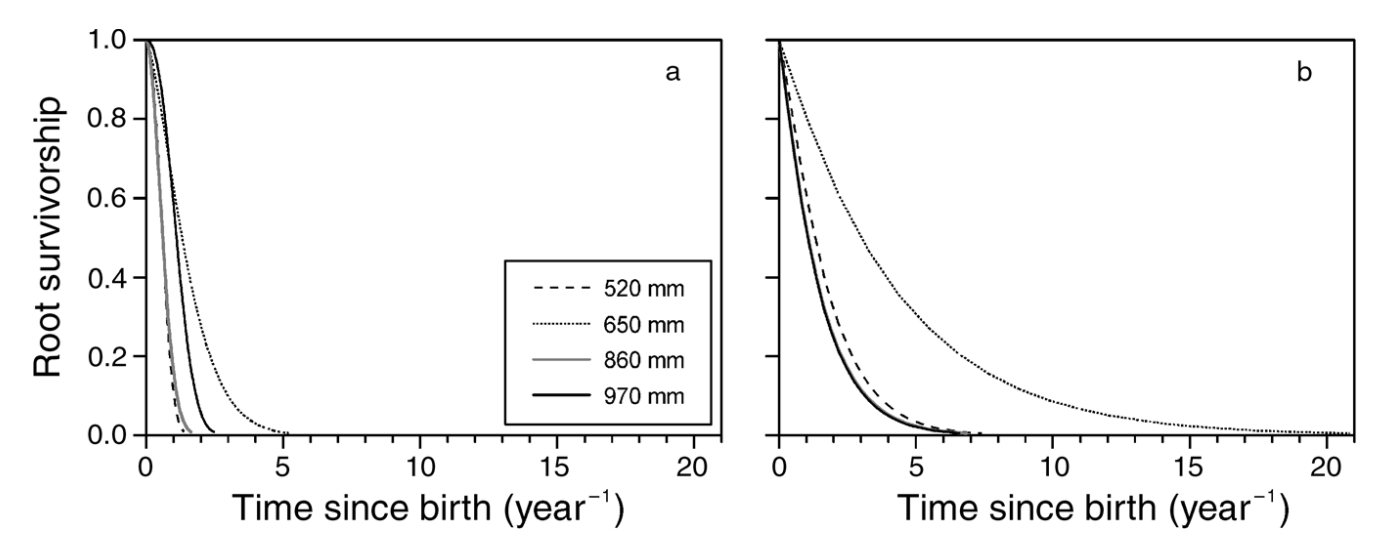
Figure 1. Root survivorship curves from mini-rhizotron data of the four populations of (a) drought-treated and (b) well-watered beech (Fagus sylvatica) trees (mean of four trees per origin and treatment), Meier & Lueschner .2008. (Image credits: Tree Physiology, 28 (2), 297–309, https://doi.org/10.1093/treephys/28.2.297)
There was 30-40% less fine root growth turnover in trees subjected to drought than those in the well-watered group. Also, species from drier habitats had a higher turnover of fine roots than the species from wetter areas. Trees shed fine roots to conserve carbon during drought, and there was a 50% reduction in the lifespan of fine roots; see Figure 1. Also, there was a ten-fold reduction in the growth rate of fine roots. There was no difference in the root structure and depth due to treatments.
In saplings, roots are concentrated in the surface soil; whereas, adult trees have a more flexible root system and will send roots deeper into the soil to search for water. Hence, saplings are more susceptible to drought.
Though there was a significant difference in fine root growth between dry and wet treatments, the difference between the four varieties was negligible. So, the scientists concluded that phenotypic plasticity was more important than genotypic variation for root growth. There is some genotypic influence but it is minor. The genetic distance was less, however, between the trees from the driest area and the wettest regions—a difference of 15%.
There was no difference in the root-to-shoot ratio due to drought in the beech trees. However, genotype did influence leaf and shoot morphology and growth.
Nutrition and Root Growth During Drought
In the South American Mediterranean regions, reforestation projects suffer from the seedling mortality of the soapbark tree, Quillaja saponaria Mol. (Quillajaceae). Commoly, these challenges arise from low soil availability and inefficient nursery and planting techniques.
Scientists wanted to test if providing higher nutrition could increase drought resistance in seedlings after planting, improving their survival. They also wanted to examine the effect of fertilization under varying water regimes.
Eight-month seedlings were grown in polyethylene bags for six months in the nursery. During this time, the seedlings were given four fertilizer treatments with 0.00, 0.45, 0.90, and 1.80 g N L-1.
After six months, the seedlings were outplanted in the fields. Half the saplings were watered and half were left unwatered.
Before planting, a few random saplings were harvested to test shoot and root parameters. The parameters measured were shoot height, stem diameter, slenderness index, shoot dry mass, and shoot to root ratio, as well as root length, volume, and dry mass.
Leaf concentrations of nitrogen, phosphorus, potassium, calcium, and magnesium were determined.
Plant height, diameter, and survival were recorded each month. Xylem water potential, chlorophyll fluorescence, and stomatal conductance were determined once every two months.
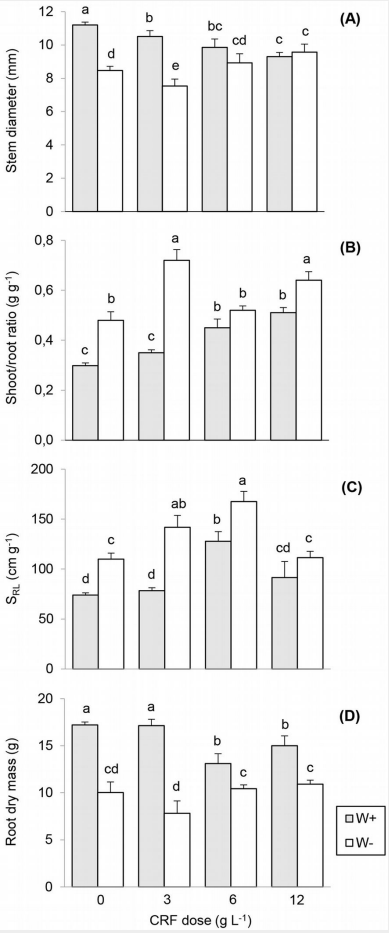
Figure 2: “Mean stem diameter (A), shoot/root ratio (B), specific root length (C) and root dry mass (D) of Quillaja saponaria seedlings grown under four nursery controlled-release fertilizer (CRF) doses two field watering regimes imposed after outplanting: watered 2 L plant-1 (W+, gray bars) and no watered (W-, white bars),” Ovalle et al. 2016. (Image credits: iForest – Biogeosciences and Forestry, Volume 9, Issue 5, Pages 758-765 (2016), doi: https://doi.org/10.3832/ifor1905-009)
Stomatal conductance was measured by the CI-340 Handheld Photosynthesis System, manufactured by CID Bio-Science Inc. This Infrared Gas Analyzer estimates stomatal conductance by determining transpiration rate as a function of leaf temperature, so readings were taken on days with similar warmth.
After one growing season, more plants were harvested, and the shoot and root parameters were estimated again.
The scientist found that adding fertilizers influenced the morphology of the plants but did not affect their chances of survival.
When plants were outplanted, the biomass allocation changed in all treatments to favor root growth. In the field, it was the unfertilized and watered plants that showed better root growth, with the lowest shoot-to-root ratio, and they had a higher stem diameter. Though they had more rootmass, it was the highly fertilized plants in the nursery that had the best root growth.
Plants that received higher nitrogen fertilization had better shoot growth after outplanting, and this was explained by higher phosphorus and nitrogen in their leaves. Both of these elements are important for shoot development.
However, their drought resistance was not improved.
In the field, watered plants had higher chlorophyll fluorescence, water potential, and higher stomatal conductance, especially the plants that received no fertilizers. So, the ecophysiology of plants was also not improved by fertilizers.
In this experiment, a higher nutrient addition in the nursery didn’t help the soapbark saplings in developing drought-resistant features later in their growth. Irrigation in the initial outplanted stages was more important.
Drought Resistance Starts From The Beginning
Saplings are more susceptible to drought, as they have not developed deep enough roots to access water, despite increased biomass allocation to roots in the sapling stage. The influence of genotype and phenotype can vary for different morphological attributes, making their influence more nuanced. Moreover, each species’ response to dryness will differ. Therefore, more research is necessary before we can make general conclusions.
—
Vijayalaxmi Kinhal
Science Writer, CID Bio-Science
Ph.D. Ecology and Environmental Science, B.Sc Agriculture
Feature image courtesy of Chris Bowman
Sources
Carbon Interactive Brief. The impacts of climate change at 1.5C, 2C and beyond. Retrieved from https://interactive.carbonbrief.org/impacts-climate-change-one-point-five-degrees-two-degrees/
Lambert, J. (2020, January 3). Climate change is bringing earlier springs, which may trigger drier summers. Science News. https://www.sciencenews.org/article/climate-change-bringing-earlier-springs-may-trigger-drier-summersScience
Meier, I.C., & Leuschner, C. (2008). Genotypic variation and phenotypic plasticity in the drought response of fine roots of European beech, Tree Physiology, 28 (2), 297–309, https://doi.org/10.1093/treephys/28.2.297
Ovalle, J.F., Arellano, E.C., Oliet, J.A., Becerra, P., & Ginocchio, R. (2016). Linking nursery nutritional status and water availability post-planting under intense summer drought: the case of a South American Mediterranean tree species. iForest – Biogeosciences and Forestry, 9 (5), 758-765. doi: https://doi.org/10.3832/ifor1905-009
Related Products
Most Popular Articles
- Transpiration in Plants: Its Importance and Applications
- Leaf Area – How & Why Measuring Leaf Area…
- How to Analyze Photosynthesis in Plants: Methods and Tools
- Plant Respiration: Its Importance and Applications
- The Forest Canopy: Structure, Roles & Measurement
- Stomatal Conductance: Functions, Measurement, and…
- Forest & Plant Canopy Analysis – Tools…
- Root Respiration: Importance and Applications
- 50 Best Universities for Plant Science
- The Importance of Leaf Area Index (LAI) in…



