Optimizing Micronutrients in Rice and Citrus using Photosynthetic Measurement

Scott Trimble
November 21, 2020 at 1:43 am | Updated March 14, 2022 at 12:09 pm | 7 min read
Several micronutrients are involved in the metabolic activities of plants, and their deficiency can affect both crop health and yield. The elements that are connected with the photosynthesis system have a substantial bearing on the biomass plants can produce. Their role in carbohydrate production, however, also allows detection of their deficiency by directly measuring photosynthetic rates. Small, portable instruments allow these measurements to be taken quickly in the field without any hassle.
The Availability of Micronutrients in Soils
As their name implies, micronutrients are needed in very small quantities by plants, but this does not mean they unimportant. They can be minerals necessary for plant growth, and their deficiency can be as detrimental to crops as that of macronutrients.
Also called trace elements, eight micronutrients are soil-derived and essential for plant growth. These are boron (B), chlorine (Cl), copper (Cu), iron (Fe), manganese (Mn), zinc (Zn), nickel (Ni), and molybdenum (Mo).
Subscribe to receive our monthly round-up of articles.
Micronutrient deficiency can occur for a variety of reasons. The parent soil material may simply not contain the micronutrients, or they are present in the soil in forms that the plant is unable to use. Other factors of the soil, like pH, water content, and compaction, can also make them unavailable to plants.
While it is not easy to predict the precise distribution of micronutrients, generally, Fe, Zn, and Mn are present in adequate amounts in acidic soils, while B is high in alkaline soils. Cu and Mo are typically found in soils with high organic matter content.
Nevertheless, intensive agriculture and high-yielding crops can deplete soils of these trace elements enough to cause physiological problems in succeeding crops, so they must be replenished with amendments.
Influence of Zn, Cu, and B on Citrus Yield
Deficiency can hit at different stages and is also an issue in both annual and perennial crops.
Orchards in Pakistan are deficient in Zn, Cu, and B. So, scientists endeavored to confirm whether these low concentrations were, in fact, affecting yield and quality of citrus.
Zn, Cu, and B are integral to enzyme activation. Other specific roles of the three micronutrients in photosynthesis are as follows:
- B is needed to transport carbohydrates and in cell development.
- Zn is necessary for chlorophyll formation and photochemical reactions.
- Cu stabilizes the chlorophyll structure and participates in electron transfer during photosynthesis.
Their deficiency can ultimately cause stunting of plants and a reduction in yield. Zn and Cu deficiency lead to chlorosis or yellowing of leaves, and B deficiency causes a reduction in flowering and fruiting.
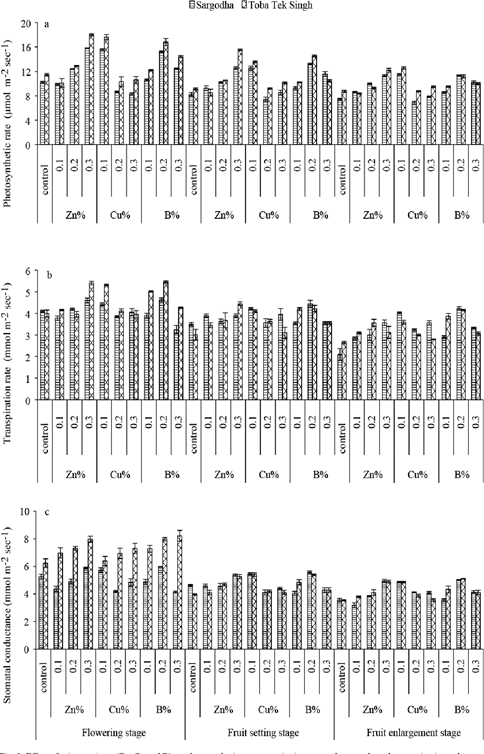
Fig. 1. “Effect of micronutrients (Zn, Cu, and B) on photosynthetic rate, transpiration rate and stomatal conductance in citrus plants at three crop stages, at Sargodha and Toba Tek Singh orchard,” Ilyas et al. 2015. (Image credits: Pak. J. Bot., 47(4): 1241-1247, 2015.)
The scientists chose two orchards of mandarin, Citrus reticulata (var. Kinnow). The control trees were sprayed with distilled water, and the three treatments with 0.1%, 0.2%, and 0.3% each of Zn, Cu, and B. Aqueous solutions of ZnSO4.7H2O, CuSO4.7H2O, and H3BO3 were given three times as foliar sprays at 15 days before bloom, fruit set, and fruit enlargement stages.
At the start and end of the experiment period, gas exchange rates—net assimilation (A), transpiration (E), and stomatal conductance (gs)—were estimated using the CI-340 Handheld Photosynthesis System.
The CI-340 is a portable infrared gas analyzer, manufactured by CID Bio-Science Inc. It is used in combination with a leaf chamber, which comes in ten sizes to fit various leaves and needles. Rapid, non-destructive measurements are possible on the spot. There are modules to control temperature, CO2, water vapor, and light intensity.
The levels of chlorophyll content were also measured before and after the experiment. In the end, fruit weight and juice volume were compared in fruit from the control and treated trees.
Supplying micronutrients improved the levels of chlorophyll and the rate of photosynthesis, transpiration, and stomatal conductance. However, the optimum level of application differed for the three elements; see Figure 1.
- Applications of Zn up to 3% improved plant physiology.
- 1% of B was optimum, but higher levels of this element were toxic and decreased gas exchanges.
- Plant physiology improved with increasing Cu concentration up to 2%, but higher levels were toxic. Toxicity of Cu also resulted in reducing the chlorophyll content.
The fruit count/tree, fruit weight, and juice volume were best in foliar applications with 0.3% Zn, 0.1% Cu, and 0.2% of B.
Scientists were able to recommend the ideal dosage of micronutrients to improve citrus yield. They also concluded that the photosynthetic system and its drop accurately predicted trends in yield decreases.
Selenium is Beneficial in Increasing Rice Yield
Selenium (Se) is more important for animals and people than it is for plants. Cereal crops grown in soils deficient in Se will have less of the element in their resulting grains and will be less nutritious. In China, Se deficiency in people has been traced back to rice, which forms a large part of the typical Chinese diet.
Selenium (Se) is not considered an essential micronutrient for plants. In fact, elemental Se is better known for its potential toxicity, but when it is converted by plant metabolism into useable compounds, it can be beneficial.
Considering its importance to people’s health, Chinese scientists wanted to find out how Se is used by the plant throughout the rice cycle, and how it is accumulated in grains and other parts of the rice plant. They also wanted to find out if Se concentrations had any effect on photosynthesis.
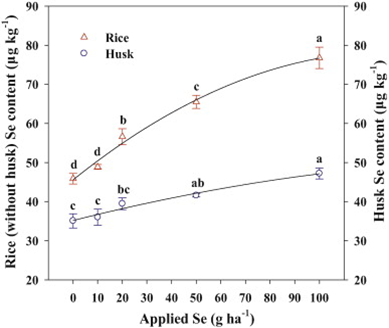
Figure 2: “Effects of Se on the rice and husk Se contents at the mature stage,” Zhang et al. 2014. (Image credits: M. Zhang et al. / Environmental and Experimental Botany 107 (2014) 39–45)
In field trials, rice was treated with five levels of Se: 0, 10, 20, 50, and 100 g Se ha−1. The soil application was in the form of selenite (Na2SeO3, AR).
Soil and plants were tested for Se at tillering, jointing, booting, flowering, and maturity. Six plants were randomly chosen, oven-dried, and analyzed for Se levels through assays.
During flowering, photosynthetic and transpiration rates were estimated using a CI-340 Handheld Photosynthesis System. Modules were used to keep the leaf temperature at 31oC, CO2 at 200ppm, and light intensity around 1200 mol m−2 s−1.
At the same time, intercellular CO2 concentration and chlorophyll fluorescence were also estimated.
The scientists found that a very narrow range of Se was beneficial for the rice plants. Levels that were lower and higher than this range were not conducive for plant growth. Application at the rate of 50 g Se ha−1 was the ideal level of Se for plant growth, but 100 g Se ha−1 was needed to produce rice with sufficient Se for people; see Figure 2.
As Se levels increased, so did its concentration in the plants. The highest level of accumulation was in the roots. Grains had the lowest, unfortunately, with shoots being intermediate. Se was used most by roots and shoots during tillering; subsequently, levels decreased, which possibly reflected decreases in soil.
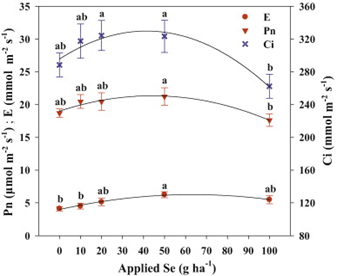
Figure 3: “Effects of Se on photosynthesis rate (Pn), transpiration rate (E) and intercellular CO2 concentration (Ci) in rice crops,” Zhang et al. 2014. (Image credits: M. Zhang et al. / Environmental and Experimental Botany 107 (2014) 39–45).
Raising concentrations up to 50 g Se ha−1 increased photosynthesis. Higher levels resulted in toxcicity and reduced photosynthesis. These changes in biomass production were mirrored in changes due to an increase in intercellular carbon and stomatal opening, as well.
Chlorophyll fluorescence also confirmed that applications up to 50 g Se ha−1 increased photosynthesis and that higher doses decreased it.
The scientists were able to see the effect of Se on the yield of rice and find the inner physiological processes that controlled it, namely the rate of photosynthesis. Levels that are toxic to plants are needed for the ideal levels of Se in grains for this cultivar, so the researchers recommended using other cultivars that were more effective at accumulating Se in the grains, as opposed to the other parts of the plant.
The Correct Dose
More is not always better. Supplementation of micronutrients should be in optimum doses, as higher concentrations can quickly turn toxic. This is true not only for beneficial elements like selenium but also for essential nutrients like copper, zinc, and boron. The photosynthetic rate can be monitored to track the impact of micronutrients on yield, as the process is sensitive to their concentrations. Using proper nutrient application and modern tools to monitor plant processes, increasing food production using existing resources is possible.
—
Tools: |
See More: |
| Pulses and Effective Drought Response
How Stress at Early Stages Affects Plants Emerging Irrigation Methods for Vineyards |
—
Vijayalaxmi Kinhal
Science Writer, CID Bio-Science
Ph.D. Ecology and Environmental Science, B.Sc Agriculture
Feature image courtesy of Dai Luo
Sources
Ilyas, A., Ashraf, M.Y., & Hussain, M., Ashraf, M., Ahmed, R., & Kamal, A. (2015). Effect of micronutrients (Zn, Cu and B) on photosynthetic and fruit yield attributes of Citrus reticulata Blanco var. Kinnow. Pak. J. Bot., 47(4): 1241-1247. Retrieved from https://pdfs.semanticscholar.org/a35d/172171366477ccc96edc496591813488f34f.pdf
Libretexts. (2019, November 19). 31.1C: Essential Nutrients for Plants. Retrieved June 23, 2020, from https://bio.libretexts.org/Bookshelves/Introductory_and_General_Biology/Book:_General_Biology_(Boundless)/31:_Soil_and_Plant_Nutrition/31.1:_Nutritional_Requirements_of_Plants/31.1C:_Essential_Nutrients_for_Plants
Mahler, R. L. (2004, Nov). Nutrients Plants Require for Growth. Retrieved from https://www.extension.uidaho.edu/publishing/pdf/CIS/CIS1124.pdf
Masunaga, T., and Marques Fong, J. D. (2018). “Chapter 11 – strategies for increasing micronutrient availability in soil for plant uptake,” in Plant Micronutrient Use Efficiency, eds M. A. Hossain, T. Kamiya, D. J. Burritt, L.-S. Phan Tran, and T. Fujiwara (Cambridge, MA: Academic Press),195–208.https://doi.org/10.1016/B978-0-12-812104-7.00013-7
Morgan, J. B. & Connolly, E. L. (2013) Plant-Soil Interactions: Nutrient Uptake. Nature Education Knowledge 4(8):2. Retrieved from https://www.nature.com/scitable/knowledge/library/plant-soil-interactions-nutrient-uptake-105289112/
Sajedi, N. A., Ardakani, M. R., Madani, H., Naderi, A., & Miransari, M. (2011). The effects of selenium and other micronutrients on the antioxidant activities and yield of corn (Zea mays L.) under drought stress. Physiology and molecular biology of plants: an international journal of functional plant biology, 17(3), 215–222. https://doi.org/10.1007/s12298-011-0067-5
Zhang, M., Tang, S., Huang, X., Zhang, F., Pang, Y., Huang, Q., & Yi, Q. (2014). Selenium uptake, dynamic changes in selenium content and its influence on photosynthesis and chlorophyll fluorescence in rice (Oryza sativa L.). Environmental and Experimental Botany, 107, 39-45. doi:10.1016/j.envexpbot.2014.05.005
Related Products
Most Popular Articles
- Transpiration in Plants: Its Importance and Applications
- Leaf Area – How & Why Measuring Leaf Area…
- How to Analyze Photosynthesis in Plants: Methods and Tools
- Plant Respiration: Its Importance and Applications
- The Forest Canopy: Structure, Roles & Measurement
- Stomatal Conductance: Functions, Measurement, and…
- Forest & Plant Canopy Analysis – Tools…
- Root Respiration: Importance and Applications
- 50 Best Universities for Plant Science
- The Importance of Leaf Area Index (LAI) in…


