Mycorrhizal fungi Associations in Various Forest Types and Climate Change

Dr. Vijayalaxmi Kinhal
July 1, 2026 at 5:47 pm | Updated July 1, 2026 at 5:47 pm | 8 min read
Key Takeaways
- Global change, such as higher carbon dioxide, warming, pests, and diseases, has common effects on mycorrhizal community composition and abundance.
- The effects of drought, forest fires, and nitrogen deposition on the mycorrhizal community differ among forest types.
- As the temperature rises, the abundance of arbuscular mycorrhizal fungi is increasing, and ectomycorrhizal fungi are decreasing in forests.
Forests cover 30% of the terrestrial surface. They are major primary producers and carbon sinks, and they influence nutrient and water cycling. Forest functioning is fluctuating due to climate change and nitrogen deposition. Global change affects microbes, including mycorrhizal fungi, which are also crucial in moderating its effects on trees and their functions. This article, primarily based on a review by Baldrian et al. (2023), explores how ongoing global changes affect mycorrhizal communities.
Types of Mycorrhizal Fungi
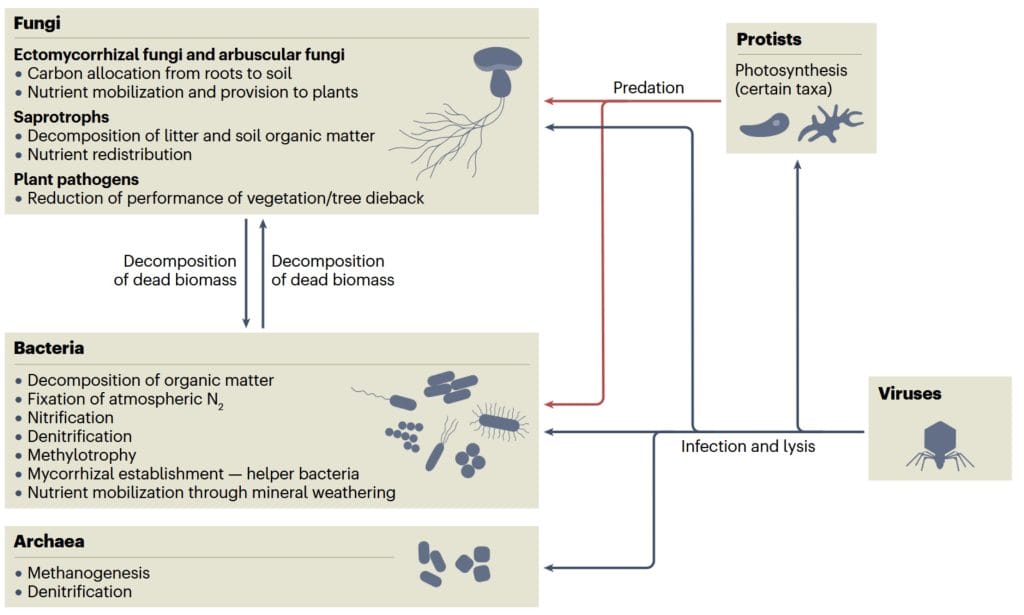
Forests host a specific soil microbiome, especially in their rhizosphere, comprising bacteria, fungi, viruses, archaea, and protists that determine ecosystem functioning.
Subscribe to receive our monthly round-up of articles.
Figure 1. Roles of the members of forest microbiomes – fungi, bacteria, archaea, protists, and viruses. Adapted from Baldrian et al. (2023). Image credit: https://doi.org/10.1038/s41579-023-00876-4
Protista are predators of fungal and bacterial cells. Archaea are restricted to specific microhabitats and are involved in methane and nitrogen cycling. The contribution of viruses to ecosystem functioning is still unclear. Bacteria that decompose plant and microbial biomass are involved in the nitrogen cycle and are responsible for atmospheric nitrogen fixation, nitrification, and denitrification. Bacteria and fungi are involved in mineral weathering and nutrient mobilization. Fungi can be saprotrophic, decomposing soil litter; pathogenic, reducing disease pressure on hosts; or symbiotic.
Mycorrhizal fungi are symbiotic and live on or inside trees’ roots, and their interactions with forest trees are crucial to tree establishment, growth, health, survival, and productivity. Mycorrhizal fungi play a crucial role in mobilizing and sequestering nitrogen and phosphorus and transporting carbon from plants to the soil. Three main types of mycorrhizae exist in forests:
Arbuscular mycorrhizal fungi (AMF): These are fungi that form mycorrhizal symbiosis with a plant host by penetrating plant root cells, where they form highly branched structures called arbuscules. The AMF belong to Glomeromycota, are associated with annual crops, and are dominant in unseasonably warm tropical regions. They are found to a small extent with temperate forest species. These fungi do compose organic matter made of biopolymers from wood, litter, and roots. Biopolymers can consist of structural compounds like lignin, cellulose, hemicellulose, and pectin from plant cell walls, peptidoglycan in bacterial biomass, or chitin in fungal matter. AMF are involved in the use of mineral forms of nitrogen and phosphorus.
Ectomycorrhizal fungi: The hyphae of this group of symbiotic fungi enclose the fine roots of plants and belong to the phyla Basidiomycota and Ascomycota. Ectomycorrhizal fungi can decompose organic matter and use organic forms of nitrogen, and are more efficient and beneficial for trees in nutrient-limited soils. Ectomycorrhizal fungi are common in cooler and drier ecosystems with lower decomposition rates, at high altitudes and latitudes, such as boreal and temperate forests.
Ericoid mycorrhizal fungi: This group of fungi forms symbiosis with some members of the plant family Ericaceae by penetrating their root hair cells and forming hyphal coils. They belong to the phyla Basidiomycota and Ascomycota and are very efficient at decomposing biopolymers. These mycorrhizal fungi are mainly found only in heathlands.
Global Change Effects on Forests
Global changes such as nitrogen deposition, warming, and elevated carbon dioxide (eCO2), directly and indirectly influence forests and their environments. Some effects of these global changes are universal. However, the importance of global change varies across forests due to differences in species diversity, climate, and dominant mycorrhizal symbioses; see Figure 2.
Baldrian et al. (2023) reviewed the trends of global changes in three forest areas – boreal, temperate, and tropical – and these are discussed below.
Forest Type Summary
| Forest type | Climate | Dominant mycorrhizal symbiosis | Main expected stressors |
| Boreal | Cold, long winters; low annual precipitation | Mostly ectomycorrhizal, with some ericoid fungi | Drought, fires, pathogens, warming, nitrogen deposition |
| Temperate | Hot summers, cold winters; moderate precipitation | Mostly ectomycorrhizal; AMF in some regions | Drought-induced mortality, fires, insects, nitrogen deposition, eCO2 |
| Tropical | Year-round warm temperatures; wet and dry seasons | Mostly arbuscular mycorrhizal; some ectomycorrhizal trees | Drought, fire severity, phosphorus limitation |
Boreal Forests
Boreal forests are the largest terrestrial ecosystems in the temperate regions of the globe. They are cold ecosystems with winters lasting over six months, average summer temperatures of 10 °C, and low annual precipitation (300-900 mm). The tree diversity is low, with ericoid shrubs in the understory. Productivity is low, allowing carbon sinks to accumulate. The dominant mycorrhizal symbiosis is ectomycorrhizal with some ericoid fungi. These ecosystems are expected to increase productivity due to CO2 fertilization, warming, and nitrogen deposition, but will suffer from drought, fires, and pathogens.
Temperate Forests
These forests have hot summers and cold winters, with temperatures ranging from -30 to 30 °C and moderate annual precipitation of 750-1,300 mm. The forests are more diverse and productive than boreal forests, but less so than in the tropics. Ectomycorrhizal symbiosis is dominant in most trees. AMF is found in some regions alone or in coexistence with ectomycorrhizal trees. Temperate forests are expected to experience negative consequences from drought-induced mortality, fires, insects, nitrogen deposition, and eCO2. Higher nitrogen deposition leads to saturation and nutrient losses, and to changes in the relative proportions of ectomycorrhizal and arbuscular trees.
Tropical Forests
Tropical regions have year-round temperatures above 20 °C but experience wet and dry seasons. Rainforests receive heavy rainfall (2,000-10,000 mm), whereas drier tropical forests receive less (above 800 mm). The tropical forest has high species diversity and productivity, with the largest above-ground carbon sinks, but lower soil carbon banks. Arbuscular mycorrhizal symbiosis is dominant, though some ectomycorrhizal trees are also present. Tropical forests will suffer mainly from drought. Nitrogen deposition increases growth, leading to phosphorus limitation.
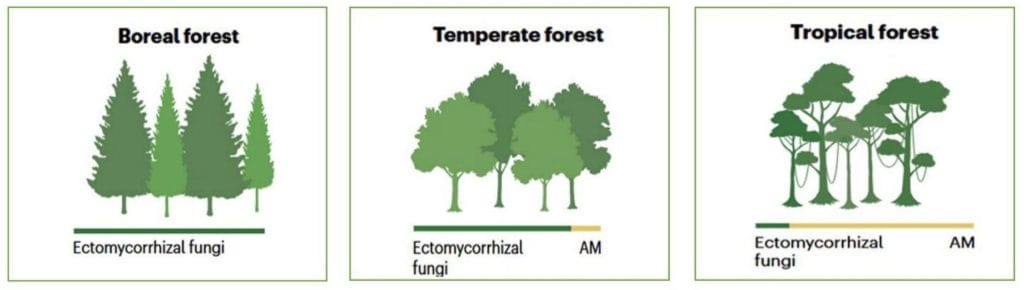
Figure 2. Dominant mycorrhizal symbioses in boreal, temperate, and tropical forests. Adapted from Baldrian et al. (2023). Image credit: https://doi.org/10.1038/s41579-023-00876-4
Differences in climate influence the species pool, the composition of tree-fungal symbioses, and nutrient cycling. Some effects of global changes on mycorrhizal symbioses are universal, but significant differences also exist.
Global Change Effects on Microbes
Some of the effects of global change on mycorrhizal symbioses that are common across the ecosystems arise from carbon dioxide fertilization, warming, pests, and diseases. The effects of drought, forest fires, and nitrogen deposition on mycorrhizal symbioses differ among forest types.
Elevated Carbon Dioxide Levels
Soil CO2 levels are usually higher than in the air, so increases in air CO2 are not expected to affect the mycorrhizal community or other microbes; changes may be restricted to Acidobacteria.
Elevated Temperature
An increase in temperature will increase the dominance of arbuscular mycorrhizal trees. AMF depend on higher temperatures and acidification. Nitrogen deposition increases soil acidification, which, along with higher temperatures, increases AMF abundance. Ectomycorrhiza occur within narrow ranges of low mean annual temperatures, so an increase in temperature reduces their abundance and species diversity. So global warming will have a disproportionately negative effect on ectomycorrhizae compared with AMF.
As the ratio of ectomycorrhiza to AMF decreases, so will their hyphal exudates, which in turn lower the spatial diversity of bacteria. More AMF will lead to faster nitrogen cycling and losses from the soil. Also, as plants grow faster due to more nitrogen, the available phosphorus levels will decrease. As temperatures rise, the vegetative phase of saprotrophic and ectomycorrhizal fungi increases, reducing the fruiting period.
Higher temperatures are thawing permafrost in boreal regions and changing fungal diversity patterns. However, tree shifts occur significantly more slowly than climate shifts; so, any increase in ectomycorrhizal fungi is sluggish.
Pests and Pathogens
Global change disturbances reduce tree activity due to windthrow, insect, and pathogen attacks. As a result, microbial numbers are lower, especially those of mycorrhizal fungi that form symbiotic relationships with trees. Their taxa, along with those of other fungi associated with dead tree biomass, will decrease in proportion to bacteria. Compared to boreal and temperate forests, which experience high pest and disease outbreaks due to increasing drought, tropical forests will be less affected by pests and pathogens due to their high tree diversity.
Drought Effects
Drought reduces net above-ground, below-ground, and total primary production worldwide. There is a corresponding reduction in shoot, root, and total tree biomass. During drought, higher pH reduces bacterial abundance and increases fungal abundance because fungal hyphae enable better water distribution within the body than bacterial cells.
Specific drought effects on mycorrhizal fungi can differ between ecosystems.
- Boreal: Drought-induced tree mortality is increasing in some boreal regions, but changes to mycorrhizal fungi are not reported.
- Temperate forests: Short-duration drought has little effect on microbes. However, as drought frequency and intensity increase, the microbial community changes. The proportion of fungi and filamentous bacteria increases at the expense of the diversity and abundance of the populations of dominant ectomycorrhizal fungi.
- Tropical forests: Contrary to expectations, tropical soil microbes are more sensitive to small changes in moisture availability, but the impact of drought on mycorrhizal fungi and other microbes remains unclear.
Forest Fires
Fires reduce trees, their activity, and microbial numbers. The effects of forest fires on mycorrhizal fungi differ significantly among the three ecosystems.
- Boreal: Forest fires are common and destroy 1% of these forests annually. Intense fires reduce organic matter, lowering carbon and nitrogen levels. As a result, microbial abundance, including that of fungi, is reduced. Bacterial diversity is not affected. The diversity of fungi, including that of ectomycorrhizal fungi, is reduced, resulting in an increase in the ratio of ericoid mycorrhizal to ectomycorrhizal fungi. Other fungi that can actively decompose organic matter are also favored, reducing carbon sinks. Post-fire microbial communities require decades to regain pre-fire community composition.
- Temperate: Increasing frequency and intensity of fires affect temperate forests, too. Decomposition of organic matter is slowed by fire but recovers faster than in boreal forests. Ectomycorrhizal fungi help their hosts acquire nitrogen, boosting the postfire relative abundance of ectomycorrhizal trees.
- Tropical: Drought is likely to increase the severity of fires in the tropics. Repeated fires create a positive feedback loop, and vegetation is unable to fully recover, with consequences for the usual microbes associated with them, namely a reduction in AMF.
Nitrogen Deposition
Nitrogen deposition effects also vary with forest types as described below.
Boreal: Since boreal forests are limited by nitrogen, higher nitrogen deposition will increase productivity. Nitrogen addition to soils increases bacterial abundance and diversity but reduces ectomycorrhizal fungi, since trees do not need them to mobilize nitrogen from organic sources.
Temperate: Higher comparative anthropogenic nitrogen deposition is seen in temperate regions. As a result, there are significant changes in the microbial community. Nitrophilic fungi that use inorganic nitrogen increase, but ectomycorrhizal fungi diversity and abundance fall because their activity is less important to their host trees. As higher temperatures favor the germination and establishment of arbuscular mycorrhizal trees in some areas of the eastern USA, there is a shift from ectomycorrhizal to AMF trees. An increase in AMF boosts nitrogen mineralization and losses from temperate forest soils.
Tropical forests: Nitrogen deposition is expected to increase productivity only in areas with nitrogen-limited soils. The ectomycorrhizal fungi are expected to decrease.
Studying Impacts on Mycorrhizal Fungi
Scientists require advanced tools to study mycorrhizal fungi in soil, such as minirhizotrons. These are standard methods because they are non-destructive, precise, and enable time-series experiments. Transparent root tubes are installed in the soil close to the subject plant/tree, allowing the root system to develop. After a few weeks, scientists can begin data collection, using root imagers, such as those offered by CID BioScience Inc. – the CI-600 In-Situ Root Imager and CI-602 Narrow Gauge Root Imager, suitable for 2 in and 2.5 in wide root tubes, respectively. The high-resolution scans taken at targeted soil depths by these imagers capture the growth and distribution of mycorrhizal fungal hyphae associated with plant roots.
Contact us at CID BioScience Inc., to learn more about our minirhizotron systems for your mycorrhizal research projects.
Sources
Baldrian, P., López-Mondéjar, R. & Kohout, P. Forest microbiome and global change. Nat Rev Microbiol 21, 487-501 (2023). https://doi.org/10.1038/s41579-023-00876-4
Clemmensen, K. E. et al. Roots and associated fungi drive long-term carbon sequestration in boreal forest. Science 339, 1615-1618 (2013).
Muys, B., Ellison, D., and Wunder, S. (n.d.). What role do forests play in the water cycle? Retrieved from https://efi.int/forestquestions/q7_en
Peterson, R. L., Piché, Y., & Plenchette, C. (1984). Mycorrhizae and their potential use in the agricultural and forestry industries. Biotechnology advances, 2(1), 101-120. https://doi.org/10.1016/0734-9750(84)90243-x
Related Products
Most Popular Articles
- Transpiration in Plants: Its Importance and Applications
- Leaf Area – How & Why Measuring Leaf Area…
- How to Analyze Photosynthesis in Plants: Methods and Tools
- Plant Respiration: Its Importance and Applications
- The Forest Canopy: Structure, Roles & Measurement
- Stomatal Conductance: Functions, Measurement, and…
- Forest & Plant Canopy Analysis – Tools…
- Root Respiration: Importance and Applications
- 50 Best Universities for Plant Science
- The Importance of Leaf Area Index (LAI) in…