How Do Stomatal Traits and Transpiration Efficiency Impact Crop Yield?
Dr. Vijayalaxmi Kinhal
June 23, 2025 at 10:16 pm | Updated June 23, 2025 at 10:16 pm | 8 min read
- Stomatal traits must meet mesophyll demand for CO2, conserve water, and maintain optimum leaf temperatures for higher transpiration efficiency.
- The stomatal traits associated with transpiration efficiency include size, density, patterning, guard cells, and responsiveness to environmental factors.
- So far, no crop breeding effort using stomatal traits has been able to prevent water loss/ transpiration without reducing photosynthesis.
Most of the gaseous exchange between plants and the environment occurs through the stomata. Stomatal conductance regulates carbon dioxide uptake for photosynthesis and water loss during transpiration, determining water use efficiency and plant productivity. Research efforts aim to balance reduced water use and increased photosynthesis to enhance transpiration efficiency by focusing on stomatal traits. Discover the key stomatal traits essential for maximizing transpiration efficiency.
Why is Stomatal Conductance in Focus?
Transpiration efficiency (TE), at the plant level, is described as the net dry matter accumulated per unit of water transpired. At the plot level, it is equivalent to water-use efficiency and represents the yield per unit of water received by plants.
Stomata make up only 3% of a leaf’s area but are responsible for 98% of carbon dioxide (CO2) uptake and water loss. Stomata thus control both photosynthetic and transpiration rates and can be key to transpiration efficiency. Stomatal conductance (gs) also boosts yield by evaporative cooling.
Subscribe to the CID Bio-Science Weekly article series.
By submitting this form, you are consenting to receive marketing emails from: . You can revoke your consent to receive emails at any time by using the SafeUnsubscribe® link, found at the bottom of every email. Emails are serviced by Constant Contact
Higher stomatal conductance is correlated with more photosynthesis and faster growth when other factors are not limiting, as more CO2 is absorbed. However, higher gs is associated with less water-use efficiency. Lower gs produce higher water-use efficiency but reduce photosynthesis and plant growth. Well-watered plants have lower stomatal conductance but can reduce photosynthesis by 20% in C3 plants and, to a lesser extent, in C4 plants, which can impact crop yield.
Water-saving stomata traits are necessary to reduce irrigation demands and prevent water stress while maintaining yield levels. The choice of water-saving characteristics will have to be customized for different regions.
Leaf stomatal traits are considered to be of greater influence than other leaf anatomical features in explaining variations in gross primary productivity (GPP). For example, in bamboo, leaf stomata contributed to 20.8% of the gross primary productivity (GPP) variation. However, the environment was also responsible for 58.3% of the variation in stomatal traits.
Research to increase transpiration efficiency has leveraged the following stomatal traits:
- Size
- Density
- Patterning
- Guard cells
- Response speed
Several new lines of crops have been developed using these approaches, which have enabled scientists to assess the resulting transpiration efficiency. The influence of anatomical and morphological traits becomes vital in determining stomatal response time and physiology, which are discussed below.
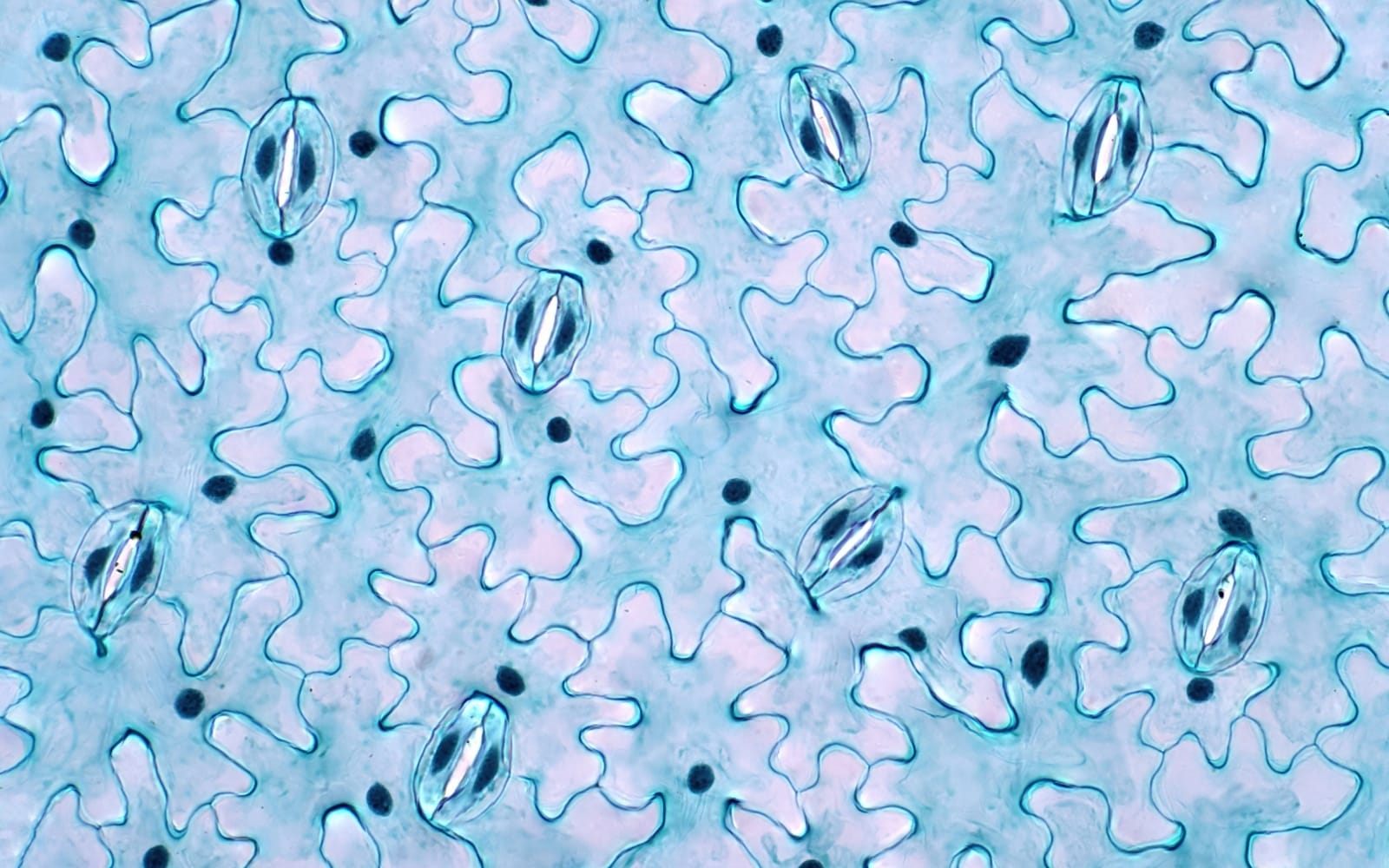
Stomatal Anatomy
![]()
Figure 1.: “The relationship between stomatal density (SD) and stomatal size (SS) in an evolutionary range of extant plants. The dotted guidelines indicate the proportion of the epidermis area allotted to stomata (A%) and, therefore, available for leaf gas exchange – A% values are indicated in the upper left corner of the figure. Lines of best-fit using non-linear regression are shown for ferns with gymnosperms (solid black line), basal angiosperms with eudicots (broken black line), monocots (broken grey line), and all species (solid grey line),” Haworth et al. 2023. (Image credits: https://doi.org/10.1016/j.scitotenv.2022.160908).
Stomatal conductance is influenced by both size and density, which are controlled by the growth environment.
Stomatal size
The stomata size and density vary across and within a species. As Figure 1 shows, there is a trend of decreasing stomata size with more evolutionarily complex plant taxa, where angiosperms have smaller stomata than conifers but larger than monocots. Stomata size can be crucial for a plant to adjust to its environment. Smaller stomata can react faster to the environment than larger stomata, and this is explained by the better surface-to-volume ratio of the guard cells, which increases solute transport and makes guard cells turgid.
Stomatal density
Gaseous conductance can be increased or decreased by manipulating stomatal density or the number of stomata per unit leaf area. Experiments aimed at reducing water use by altering stomatal density began in the 1970s. The approach was to increase conductance with higher stomatal density to boost the plant growth rate. However, the studies showed that the effect of density on transpiration is complex. Modifications in functions can counterbalance changes in the physical anatomy of stomata.
Hence, increasing density did not enhance CO2 uptake and growth; instead, it was the water status that was more significantly affected. Increasing density didn’t help, as there was less space for CO2 movement in the mesophyll. In some cases, such as osmotic stress and salinity, a lower stomatal density is more favorable for conserving available water. For example, wild barley genotypes have less stomata density than cultivated varieties but have lower productivity. Even among cultivated varieties, stomatal density reduces by 16% due to water or salinity stress. The same trend in stomatal density is also seen between wild and cultivated rice.
In one experiment on Arabidopsis thaliana, boosting stomatal density by 2-3 times that of wild type resulted in a 30% hike in photosynthesis but reduced water-use efficiency by 50%.
However, in forests, stomatal density was positively correlated and explained 51% of the variation in productivity.
Relationship between size and density
In general, stomata size and density are inversely related. When the stomata size is larger, the density is lower. The interactions between stomata size and density will affect the response of stomatal conductance to the environment.
Low densities of larger stomata exhibit a slower response to environmental changes. However, higher densities of smaller stomata do not always result in a rapid response. Other environmental factors can also be crucial. Plants with lower density and larger stomata can exhibit lower transpiration rates and higher growth rates and biomass accumulation under optimal conditions.
Stomatal Patterning
The patterning or spacing between stomata becomes important as density increases. During clustering, CO2 diffusion is restricted, which reduces photosynthesis. Usually, stomata have a “one-cell-spacing” to avoid this problem, but crop breeding sometimes produces mutants that break this rule and have lower productivity.
Clustering of stomata also results in uneven distribution of CO2. The leaf area without stomata above mesophyll had less CO2 concentrations and photosynthesis, suggesting that lateral movement of gas is not enough to reach places in the leaf without stomata.
Guard Cells
The shape and size of guard cells are also critical stomatal traits that influence their function. There are two common shapes: the kidney-shaped guard cells found in most plants and the ‘dumbbell’-shaped guard cells found in the monocots Poaceae, as shown in Figure 2.
![]()
Figure 2.: “Example stomatal complexes: a) kidney-shaped stomatal complex, and; b) dumbbell-shaped stomatal complex. GCW = guard cell width; GCL = guard cell length; SPW = stomatal pore width; SPL = stomatal pore length. Stomatal size (SS) is calculated as SS = GCL ∗ (2 ∗ GCW),” Haworth et al. 2023. (Image credits: https://doi.org/10.1016/j.scitotenv.2022.160908).
Kidney-shaped cells are larger, and the dumbbell guard cells are smaller. Hence, the dumbbell-shaped guard cells can rapidly alter turgor through ion movement across the cell membrane and respond more quickly to environmental changes. The response of dumbbell guard cells is faster in C4 plants than in C3 plants. Across species with kidney-shaped guard cells, the stomatal conductance decreases with higher density, but this relationship is not strong in dumbbell-shaped guard cells.
Some scientists consider manipulating guard cell ion movement as important as the physical traits of stomata. Stomatal response to the environment is slower by many orders of magnitude compared to changes in ion transport activity and guard cell membrane voltage.
The presence or absence of subsidiary cells is also crucial. The dumbbell-shaped guard cells in grasses (Poaceae) have subsidiary cells that maximize stomatal aperture due to a quick switch of turgor pressure from subsidiary to guard cells. Several grain crops that belong to Poaceae have benefited from the guard-subsidiary cell complex and optimal patterning to give faster stomatal responses to their environment.
Stomatal Response Speed
The external environment and internal cues influence stomatal opening and closing, and species can vary in sensitivity and responsiveness. Stomata are open when exposed to external light and high temperature, and internal CO2 and vapor pressure deficit (VPD) are low. Stomata are closed due to shade or darkness, as well as high internal CO2 and VPD levels. In nature, these factors can co-occur, and the stomata response is hierarchical. CO2 and light are related to CO2 demand in the mesophyll for photosynthesis, and VPD and high temperature are associated with the water status of the plant.
Photosynthesis rate adjustment to light and CO2 changes are faster and can occur in seconds to minutes. However, stomatal conductance in response to VPD and temperature can take minutes to hours. Therefore, the photosynthesis rate is reduced more rapidly in shade, but stomatal conductance can still occur, resulting in water loss and a decrease in transpiration efficiency. During light recovery after shade, photosynthesis capacity is revived instantly, but stomata take longer to open and let in CO2 for the process to occur. The speed of response in photosynthetic rate and stomatal conductance varies among species. There is also a difference between species in responsiveness to sun and shade flecks.
![]()
Figure 2: “Temporal responses of (a) stomatal conductance gs and (b) net CO2 assimilation A in Vicia faba (blue dots) and Avena sativa (red dots) to a step change increase followed by a decrease in irradiance. Values are an average of four replicates, and the colored shading represents plus/minus SE. The shaded and white areas represent light intensities of 100 μmol m−2 s−1 and 1000 μmol m−2 s−1, respectively. Data redrawn from McAusland et al. (2016),” Lawson and Vialet-Chabrand (2018). (Image credits: https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.15330)
Crops grown in monoculture experience sun and shade flecks due to clouds and sun angle, as well as shade from neighboring plants, and will experience fast changes in photosynthesis that must be considered. Stomata close faster in shade than they open in light. During light recovery after shade, photosynthesis capacity is revived instantly, but stomata take longer to open and let in CO2 for the process to occur.
Suppose stomatal conductance responds more quickly to environmental changes during periods of no photosynthesis. In that case, water loss can be reduced to enhance transpiration efficiency, or it can open more rapidly to support greater carbon fixation for biomass accumulation.
The photosynthesis rate can be affected by up to 10% due to the responsiveness of stomatal conductance.
No single optimal approach exists to improve transpiration efficiency by utilizing stomatal traits. Species-specific anatomy and physiology exhibit trade-offs, exemplified by the interactions between size and density. Most studies have focused on stomatal size and density; however, the results indicate that this approach is not the most effective solution for improving transpiration efficiency. Only a few studies exist on stomatal responsiveness and the mechanisms controlling its closing and opening speed, which could be essential for daily alteration of CO2 uptake and water loss.
For more research on this critical topic, scientists can use tools like those offered by CID Bio-Science Inc. – the CI-340 Handheld Photosynthesis System. It can measure stomatal conductance, photosynthesis, and transpiration simultaneously on-site in real-time. It is a small, portable precision tool based on Infrared Gas Analysis.
Contact CID Bio-Science Inc. to learn more about the CI-340 Handheld Photosynthesis System and how it can help in your research on transpiration efficiency.
Sources
Albasha, R., & Bartlett, M. K. (2024). The stomatal traits that conserve water without compromising grapevine carbon gain depend on climate change severity and wine-growing region. Agricultural and Forest Meteorology, 347, 109892.
Guo, W., Cherubini, P., Zhang, J., Li, M. H., & Qi, L. (2023). Leaf stomatal traits rather than anatomical traits regulate gross primary productivity of moso bamboo (Phyllostachys edulis) stands. Frontiers in plant science, 14, 1117564. https://doi.org/10.3389/fpls.2023.1117564
Hasanuzzaman, M., Zhou, M., & Shabala, S. (2023). How Does Stomatal Density and Residual Transpiration Contribute to Osmotic Stress Tolerance? Plants, 12(3), 494. https://doi.org/10.3390/plants12030494
Haworth, M., Marino, G., Materassi, A., Raschi, A., Scutt, C. P., & Centritto, M. (2023). The functional significance of the stomatal size to density relationship: Interaction with atmospheric [CO2] and role in plant physiological behaviour. Science of the Total Environment, 863, 160908.
Lawson, T., & Blatt, M. R. (2014). Stomatal size, speed, and responsiveness impact on photosynthesis and water use efficiency. Plant physiology, 164(4), 1556–1570. https://doi.org/10.1104/pp.114.237107
Lawson, T., & Vialet‐Chabrand, S. (2019). Speedy stomata, photosynthesis and plant water use efficiency. New Phytologist, 221(1), 93-98.
Wang, R., Yu, G., He, N., Wang, Q., Zhao, N., Xu, Z., et al. (2015). Latitudinal variation of leaf stomatal traits from species to community level in forests: linkage with ecosystem productivity. Sci. Rep. 5 (1), 1–11. doi: 10.1038/srep14454
Related Products
Most Popular Articles
- Transpiration in Plants: Its Importance and Applications
- Leaf Area – How & Why Measuring Leaf Area…
- How to Analyze Photosynthesis in Plants: Methods and Tools
- Plant Respiration: Its Importance and Applications
- The Forest Canopy: Structure, Roles & Measurement
- Stomatal Conductance: Functions, Measurement, and…
- Forest & Plant Canopy Analysis – Tools…
- Root Respiration: Importance and Applications
- The Importance of Leaf Area Index (LAI) in…
- 50 Best Universities for Plant Science



